ریاضیات جذابیت جنسی
مجله زیست شناسی جلد 9 , شماره مقاله: 18 ( 2010 ) به این مقاله استناد کنید
خلاصه
لوله های گرده به دنبال جاذبه هایی هستند که توسط تخمک ها ترشح می شوند. در مقالهای اخیر در BMC Plant Biology ، استیومن و همکارانش پارامترهای این جاذبه را اندازهگیری کردهاند و از آنها برای کالیبره کردن یک مدل ریاضی استفاده کردهاند که فرآیند را بازتولید میکند و پیشبینیهایی را در مورد ماهیت جذبکننده ماده و مکانیسمهای پاسخ نر ممکن میسازد.
مقاله پژوهشی را ببینید: http://www.biomedcentral.com/1471-2229/10/32
داروین از موفقیت سریع گیاهان گلدار در تکامل به عنوان یک راز نفرت انگیز یاد کرد. بخش عمده ای از این موفقیت به روش های عجیب و غریب تولید مثل جنسی که آنها تکامل یافته اند متکی است. حقایقی که ما را خوشحال می کند و کنجکاوی ما را در مورد رفتار جنسی حیوانات برمی انگیزد، در واقع جوهر بسیاری از رمان ها - جاذبه، فریب، شیمی، رقابت مردانه، انتخاب زن، سقط جنین، مرگ و قربانی کردن - هر بار درست زیر بینی ما اتفاق می افتد. ما از بوی خوب یک گل قدردانی می کنیم.
عاشقانی که معمولاً با چشم قابل مشاهده نیستند، گامتوفیت های نر و ماده، دانه گرده و کیسه جنین هستند. طرح در داخل اندام های تناسلی زن رخ می دهد که در مجموع به عنوان مادگی شناخته می شود (شکل 1 ). عمل نهایی شامل جذب غیرقابل مقاومت به تخمک از طریق مولکول های خاص است که به ناچار منجر به فداکاری نوع دوستانه سلول لوله گرده می شود که منفجر می شود تا سلول های اسپرم را به کیسه جنین برساند.
شکل 1
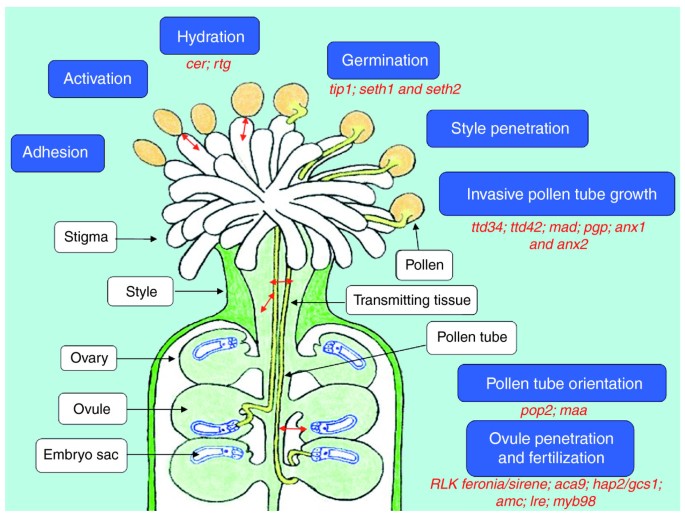
آناتومی تولید مثل جنسی در آرابیدوپسیس . یک مادگی با گرده در مراحل مختلف گرده افشانی نشان داده شده است. در یک گرده افشانی معمولی سازگار، گرده به کلاله میچسبد و روی آن جوانه میزند و یک لوله گرده تولید میکند. لوله از طریق بافت های ماده به سمت تخمدان رشد می کند، جایی که باید مسیر رشد خود را تنظیم کند تا یک تخمک پیدا کند و سپس دوباره بچرخد تا وارد میکروپیل شود و به کیسه جنین نفوذ کند. جعبه های سفید اندام های اصلی را نشان می دهد که در این فعل و انفعالات شرکت می کنند. جعبه های آبی نشان دهنده فرآیندهای اصلی هستند (از چسبندگی در بالا سمت چپ تا لقاح در پایین). متن قرمز نشان دهنده نماینده Arabidopsis استجهش یافته هایی که به هر طریقی بر این فرآیندها تأثیر می گذارند. فلش های قرمز برهمکنش اصلی سلول-سلول بین گامتوفیت نر (دانه و لوله گرده) و اندام های مختلف ماده و گامتوفیت (کیسه جنین) را نشان می دهد. در آرابیدوپسیس کل فرآیند، از چسبندگی گرده تا لقاح، حدود 4 تا 8 ساعت طول می کشد.
از زمان لینه، این فرآیند زیست شناسان را به خود جلب کرده است، و اگرچه اساس بنیادی تعاملات در قرن نوزدهم توسط محققانی مانند رابرت براون، جووانی باتیستا آمیسی، داروین و سرگئی ناواشین [ 1 ] درک شد، اما ماهیت مولکولی این جاذبه کشنده اکنون درک می شود [ 2 ]. ریاضیات در حال حاضر برای بررسی این مسئله برای اولین بار در مقاله ای نوآور که اخیراً در BMC Plant Biology [ 3 ] منتشر شده است، استفاده می شود. در این مطالعه، استیومن و همکارانش از نیمه تنی استفاده کردندسیستمی برای تعیین کمیت بهتر ماهیت و محدوده جاذبه های تخمک. مهمتر از آن، آنها یک مدل ریاضی تصادفی از تحرک کل سلول را برای رشد لوله گرده اعمال کردند. این نوع مدل قبلاً با موفقیت برای توصیف کموتاکسی در سلولهای یوکاریوتی مختلف مانند لکوسیتها و لیستریا استفاده شده است. هنگامی که با داده های تجربی جدید کالیبره شد، این مدل پیش بینی هایی را در مورد اندازه و ویژگی های فیزیکی جاذبه انجام داد. نتایج نشان میدهد که ویژگیهای رشد مشاهدهشده تا حد زیادی بازده جذب تخمکها را افزایش میدهد.
آناتومی پیچیده یک جاذبه کشنده
بیشتر زیباییهایی که گلها ممکن است از طریق ترکیب رنگها، شکلها و رایحههایشان برای ما داشته باشند، از دیدگاه رشد فقط یک هدف دارند: جذب حشرات و سایر حیوانات از طریق فریب برای انجام گرده افشانی. تولید مثل زمانی اتفاق میافتد که یک دانه گرده روی سطح پذیرای مادگی، کلاله (شکل 1 ) قرار میگیرد، چه از طریق یک رابطه پیچیده بین حشرات و ارکیدهها یا از طریق پراکندگی صرف گردههای علف توسط باد به آنجا برسد. در آنجا، کلمه "شیمی" به معنای لغوی و همچنین معنایی استعاری است، زیرا پیوند بین لایه های خارجی تخصصی دیواره سلولی گرده و سلول های کلاله بیرونی قوی تر از قوی ترین ابرچسب صنعتی محاسبه شده است. 1]. بسیاری از گیرندهها و لیگاندها برای اطمینان از تشخیص مناسب در محل قرار میگیرند، و در صورت سازگاری، سلولهای کلاله دانههای گرده بسیار کمآب را با تأمین آب و مواد مغذی به آنها پرورش میدهند و اجازه جوانهزنی را میدهند. لوله گرده سپس از دانه گرده در یک مثال شدید از رشد سلولی پلاریزه و آپیکال که از مجموعه ای غیرمعمول از ویژگی های سلولی ناشی می شود، رشد می کند [ 4 ]. لوله گرده یکی از سریعترین سلولهای در حال رشد در طبیعت است و رونویسی بسیار تخصصی را برای سیگنالدهی و ارتباطات سلولی رمزگذاری میکند، که آنها را به ماشینهای ادراک محرک کارآمد تبدیل میکند [ 1 ]. اما اینکه چگونه و چرا این فعل و انفعالات سلول-سلول انجام می شود و لوله گرده را با دقت بسیار دقیقی برای هدف قرار دادن ورودی تخمک - میکروپیل - هدایت می کند، هنوز محل بحث است.
این رویدادها همگی در اعماق بافتهای مادگی اتفاق میافتند و اخیراً امکان تصویربرداری مستقیم از آنها با استفاده از میکروسکوپ دو فوتونی وجود داشته است [ 5 ]. در دهههای پس از کار پیشگام Rosen، Mascarenhas و دیگران، شواهدی جمعآوری شده است که احتمالاً ترکیبی از مولکولهای شیمیگردان برای برخی از مراحل مورد نیاز است. با این حال، در بیشتر مسیر (از چسبندگی تا ورود به تخمدان؛ شکل 1 )، آرایش آناتومیکی بافتها به نظر میرسد برای ایجاد آزادی محدودی برای رشد لولهها از نظر مکانیکی کافی باشد. ویژگیهای فیزیکی و شیمیایی بافتهایی که با لولههای گرده تماس مستقیم دارند (لیپیدها، آب، پروتئینهای گلیکوزیله و غیره) بقیه سیگنالها را ارائه میکنند [ 1 ، 6 ].]. با ظهور ژنتیک، تعدادی از صفحات در Arabidopsis جهشها را برای اکثر مراحل این فاز به اصطلاح پروگامیک تولید مثل جدا کردند (لیست نمایندهای از جهشیافتهها برای هر مرحله به رنگ قرمز در شکل 1 نشان داده شده است ). برخی از این جهشیافتهها شواهدی برای هدفگیری دوربرد ارائه کردهاند، با سیگنالهایی که شاید تا 500 میکرومتر رخ دهد. فهرست ژنهای درگیر احتمالاً بهطور قابلتوجهی بزرگتر میشود زیرا روشهای غربالگری جدید دهها جهش خاص مردانه و زنانه را بهویژه در Arabidopsis نشان میدهند [ 7 ]، اما تاکنون هیچیک ما را به مولکول(های) کموتاکسی خاص نزدیک نکرده است.
از شیب ها و تخلیه های انفجاری
به اندازه کافی عجیب، پاسخ به سوال هویت مولکول کموتاکسی از جنین شناسی تجربی کلاسیک به دست آمد. برای مدت طولانی، تیم های مختلف به دلایل اساسی یا کاربردی، طرح های لقاح نیمه مصنوعی را توسعه داده اند [ 6 ]. در اکثر این طرحها، گردهها اجازه دارند در کلاله جوانه بزنند، اما سبک برداشته میشود و لولههای گرده اجازه رشد خارج از سبک را به یک محیط مصنوعی میدهند. این روش نیمه تنی اخیراً با تخمک های جدا شده ترکیب شده است تا این فرضیه را آزمایش کند که آیا سیستم برای ایجاد جاذبه برای لوله گرده کافی است یا خیر. این امر وجود گرادیان های جذب کننده ترشح شده از تخمک ها را تایید می کند. چنین سیستمی برای اولین بار در گیاه ساکولنت کار می کندGasteria [ 8 ] و اخیراً در گونه های مدل Arabidopsis [ 9 ]، سوسن لیلیوم [ 10 ] و ذرت [ 11 ]. اما هیچ یک از اینها به اندازه سیستم توسعه یافته توسط هیگاشیاما و همکارانش با استفاده از گل جناغی، Torenia fournieri [ 6 ] قدرتمند و آموزنده نبوده است. این نویسندگان با شروع از مشاهدات تشریحی که در این گونه گامتوفیت ماده - کیسه جنینی - برهنه و بدون هیچ گونه بافت اطراف در معرض دید قرار گرفته است [ 6 ]] مجموعه ای از آزمایشات را توسعه داد که برای اولین بار تخلیه انفجاری لوله گرده را در داخل تخمک نشان داد و سلول های هم افزایی (شکل 2 ) را به عنوان منبع سیگنال جذب کننده منتشر کننده شناسایی کرد [ 6 ]. در یک تور بینیروی از پروتئومیکس، همین نویسندگان اخیراً اولین پروتئینهایی را جدا کردند که نشان داده شده بود به طور خاص در سیگنال جذب نقش دارند: پپتیدهای کوچک، شبیه دیفنسین و غنی از سیستئین به نام LUREs [ 12 ]. اکنون به نظر می رسد مدلی از جذب لوله گرده با انتشار مولکول ها از میکروپیل به خوبی ایجاد شده است (شکل 2a ). اگر بتوان مقایسه ها را از میدان حیوانی انجام داد [ 11]، انتظار میرود که بسیاری از کلاسهای پروتئین توصیف شوند، زیرا تلاش پروتئومیکس تازه شروع شده است و بسیاری از تلاشهای غربالگری در حال انجام است [ 7 ].
شکل 2
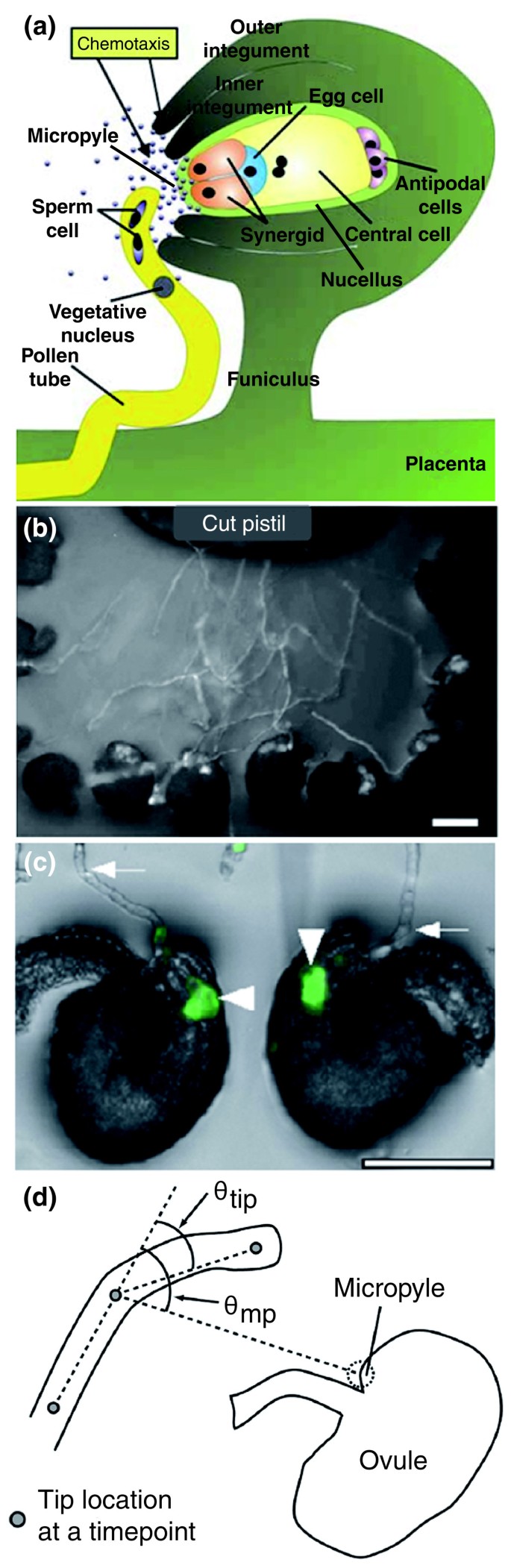
جذب تخمک و کموتاکسی رشد لوله گرده (الف) هدایت لوله گرده قبل از لقاح مضاعف در گیاهان گلدار است. یک لوله گرده حامل دو سلول اسپرم از جفت خارج می شود تا در امتداد فونیکولوس (پای تخمک) به میکروپیل (ورودی تخمک) به دنبال شیب های ایجاد شده توسط بافت های مادری تخمک و توسط گامتوفیت ماده رشد کند. کیسه جنینی حاوی دستگاه تخمک (سلول تخمک و دو سلول هم افزایی)، سلول مرکزی با دو هسته قطبی و سه سلول پادپای است. معمولاً توسط یک بافت حمایتی - هسته - و دو لایه بافت محافظ - پوشش داخلی و خارجی احاطه شده است. در تورنیاهسته متلاشی شده و یک دستگاه تخم برهنه در ناحیه میکروپیلار ایجاد می کند. اقتباس از [ 11 ]. (ب، ج) سیستم رشد نیمه تنی در آرابیدوپسیس. گرده در کلاله جوانه می زند، اما سبک برش داده می شود (بالا در (b)) و با تخمک های جدا شده (پایین در (b)) کشت می شود. هنگامی که از سبک خارج می شوند، لوله های گرده در سطح یک محیط نیمه جامد آگار رشد می کنند و در نهایت میکروپیل تخمک ها را هدف قرار می دهند (c). اگر نفوذ حاصل شود، محتویات لوله ها در داخل یک هم افزایی تخلیه می شود. اگر سیستم با لوله های گرده (فلش های در (c)) که با پروتئین فلورسنت سبز برچسب گذاری شده اند انجام شود، لحظه لقاح با فلورسانس قابل مشاهده است (سر پیکان ها در (c))، و تخمک ها را می توان از نظر جذب موفقیت آمیز نمره گذاری کرد. میله های مقیاس نشان دهنده 100 میلی متر است. اقتباس از [ 9 ]. (د) تصویری از زوایای مورد استفاده در تجزیه و تحلیل چرخش لوله گرده ساخته شده توسط Stewman و همکاران. [ 3]. این زوایا نشان می دهد که لوله گرده چقدر باید بچرخد تا مستقیم ترین مسیر را به سمت میکروپیل (q mp ) طی کند و جهت جدیدی را که توسط لوله گرده در پاسخ به گرادیان انتخاب شده است ( نوک q ) توصیف می کند. سپس این دادههای کمی برای دورههای جوجهکشی مختلف جمعآوری شدند تا ماهیت و اثر گرادیان تولید شده توسط انتشار یک ماده جاذب از میکروپیل تخمک را استنتاج کنند.
جذب از طریق شیب: در جایی که لیگاند وجود دارد باید گیرنده ای وجود داشته باشد
توسعه یک سیستم نیمه تنی مناسب لقاح برای آرابیدوپسیس (شکل 2b، c ) نه تنها استفاده از ژنتیک را امکان پذیر کرده است، بلکه برای بررسی ماهیت فیزیکی مولکول منتشر کننده در آرابیدوپسیس ، مانند تورنیا ، استفاده شده است. [ 8 ]. استیومن و همکاران [ 3 ] اکنون زوایای انحنای رشد لوله های گرده تورنیا را به دقت تجزیه و تحلیل کرده اند (شکل 2dدر شرایط آزمایشی مختلف، یعنی با زمانهای مختلف جوجهکشی تخمکها (احتمالاً مربوط به سطوح مختلف یک گرادیان ایستاده است)، و بنابراین میتواند پارامترهای کمی مهم فرآیند جذب را تعیین کند. اول، آنها دریافتند که عمل گرادیان می تواند فاصله ای بین 100 تا 150 میلی متر [ 3 ] را گسترش دهد، فاصله ای قابل توجه بیشتر از آنچه قبلاً تصور می شد [ 6 ، 9 ] یا به طور تجربی با گرادیان های مصنوعی مولکول های جدا شده آزمایش شده بود [ 2 ، 12]. این تخمین فاصله احتمالاً به این معنی است که انواع مختلفی از مولکولها با دامنهها و عملکردهای مختلف با هم جمع میشوند تا واکنش بیولوژیکی را ایجاد کنند. شکی وجود ندارد که ویژگیهای گونهای باید توسط پروتئینهای کدگذاری شده با ژن مشخص شود، و اینکه LUREهای جدا شده و پروتئین دستگاه تخم مرغ Zea mays 1 (ZmEA1) هر دو اثرات گرمسیری مثبت ایجاد میکنند. با این حال، به نظر میرسد شواهد بسیاری دیگر از مواد شیمیایی غیر اختصاصی، کوچک و قابل انتشار، مانند یونها یا حتی گاز سیگنالدهنده اکسید نیتریک (NO) را دخیل میدانند [ 10 ]]. در واقع، دخالت NO تعجب آور نیست، زیرا به نظر می رسد رفتارهای مختلف نشان داده شده توسط لوله های گرده زمانی که غلظت NO مختل می شود، نشان دهنده کند شدن رشد لوله گرده با نزدیک شدن به منبع انتشار است. این اثرات با آنچه در مدل Stewman و همکاران [ 3 ] یافت می شود، مطابقت دارد.
سادگی ظاهری رشد لوله گرده منجر به رویکردهای مختلفی برای مدلسازی ریاضی ویژگیهای اصلی آن میشود. لوله های گرده با موفقیت از طریق روش های مکانیکی یا هندسی توصیف شده اند [ 13 ]. با نگاه از زاویه ای متفاوت، برجستگی سیستم های سیگنال دهی رشد لوله گرده اخیراً با این فرض مدل سازی شده است که گیرنده های پروتئین چسبنده حساس به فاکتور N -ethylmaleimide محلول (SNAREs) و سیگنال دهی کوچک GTPase عوامل اصلی رشد هستند [ 14 ].
استیومن و همکاران [ 3 ] گامی فراتر برداشت و بر روی ویژگیهای رسمی سیستم تمرکز کرد، همانطور که از پارامترهای جنبشی دقیقاً از سیستم نیمه تنی مشتق شده است. مشکل سنجش گرادیان پیش پا افتاده نیست و نویسندگان [ 3] استراتژی ساخت بر روی مدلهای حرکتی تمام سلولی تصادفی را دنبال میکند، که اساساً فرض میکنند که لولههای گرده میتوانند تفاوتی در کسر گیرندههای متصل به یک جاذب را حس کنند و رشد خود را بسته به این بخش تغییر دهند. یکی از پیشبینیهای این مدل این بود که نرخ رشد آهستهتر در یک گرادیان ثابت جذبکننده (که فرض میشود از انتشار همسانگرد جذب کننده از تخمک وجود دارد) توانایی لولههای گرده را برای هدفگیری موفقیتآمیز تخمکها به میزان زیادی افزایش میدهد. آنها می توانند با آزمایش های خود اعتبار آماری را تأیید کنند [ 3 ]. علاوه بر این، مدل الگوهای مشاهدهشده رشد تصادفی و مستقیم مشاهدهشده در طول رشد لولههای گرده در شرایط آزمایشگاهی را توصیف میکند.
اگرچه فرض یک توصیف ریاضی کاملاً رسمی از هر پدیده بیولوژیکی ممکن است منجر به توصیفات کاملاً پدیدارشناختی با ارزش تجربی محدود شود، پیچیدگی سیستم و تعداد اجزای مختلف سلولی که به نظر میرسد برای رشد اساسی هستند [ 4 ] این رویکردها را بسیار بالا میبرد. آموزنده یا حتی اساسی برای درک مبانی مکانیکی پاسخ ماکروسکوپی سیستم. به عنوان مثال، این مدل [ 3] ممکن است به ما کمک کند تا بفهمیم چگونه لولههای گرده نشانههای هدایت خارجی را با گرادیانهای یون درون سلولی یا سایر مکانیسمهای هدایت سلولی شناخته شده مرتبط میکنند. علاوه بر این، مدل فرض می کند که حداقل دو تکه گیرنده وجود دارد که در لوله گرده از هم جدا شده اند. اگرچه این فرض چیزی در مورد گیرنده ها نمی گوید، اما نشان می دهد که یک مدل حداقلی از یک حسگر با موقعیت نقطه آپیکال دقیق کار نخواهد کرد. در عوض، از یک غشاء یا سیتوزولی جداسازی فضایی یا گیرنده ای که در عرض (حداقل) قطر لوله کشیده می شود، حمایت می کند. این مدل همچنین فرض میکند که تغییر غلظت در نوک لوله بسیار کمتر از میانگین غلظت در نوک لوله است. علیرغم اینکه تحت تأثیر محدودیت مدلسازی دو بعدی انجام شده [ 3]، این یک پیشبینی کمی قوی است که میتواند هنگام جستجوی مولکولهای جدیدی که ممکن است با مشخصات مطابقت داشته باشد تأیید شود. و چگونه کند شدن لوله ها در نزدیکی میکروپیل اتفاق می افتد؟ به عنوان مثال، NO برای کاهش سرعت رشد شناخته شده است [ 10 ]، و فرمول بندی مدل در واقع اجازه می دهد تا تعاملات چند عاملی بر روند رشد تأثیر بگذارد. نکته مهم این است که این فرمول ریاضی ممکن است به تبعیض از اثرات مختلف در طول روش های تجربی بر اساس انحراف زاویه رشد و/یا رابطه نرخ رشد با هدف گیری موفق اجازه دهد.
مانند بسیاری از رویکردهای ریاضی دیگر به رفتارهای پیچیده بیولوژیکی، این مدل جدید از Stewman و همکاران. [ 3 ] سوالات بیشتری نسبت به پاسخ ایجاد می کند. اما این واقعیت که رویکردهای جدید به توصیف تجربی دقیق سیستم کمک میکنند [ 2 ، 11 ، 12 ] ممکن است مدلسازی ریاضی را به ابزاری مهم برای آزمایش و انتخاب مولکولهای کاندید تبدیل کند که ممکن است با مشخصات بیولوژیکی in vivo مرحله نهایی گیاه مطابقت داشته باشد. جاذبه جنسی
منبع
https://jbiol.biomedcentral.com/articles/10.1186/jbiol233
 در این وبلاگ به ریاضیات و کاربردهای آن و تحقیقات در آنها پرداخته می شود. مطالب در این وبلاگ ترجمه سطحی و اولیه است و کامل نیست.در صورتی سوال یا نظری در زمینه ریاضیات دارید مطرح نمایید .در صورت امکان به آن می پردازم. من دوست دارم برای یافتن پاسخ به سوالات و حل پروژه های علمی با دیگران همکاری نمایم.در صورتی که شما هم بامن هم عقیده هستید با من تماس بگیرید.
در این وبلاگ به ریاضیات و کاربردهای آن و تحقیقات در آنها پرداخته می شود. مطالب در این وبلاگ ترجمه سطحی و اولیه است و کامل نیست.در صورتی سوال یا نظری در زمینه ریاضیات دارید مطرح نمایید .در صورت امکان به آن می پردازم. من دوست دارم برای یافتن پاسخ به سوالات و حل پروژه های علمی با دیگران همکاری نمایم.در صورتی که شما هم بامن هم عقیده هستید با من تماس بگیرید.